傕偆堦搙丄僙儖儘乕僗扨寢徎偵偮偄偰
丂暿復偱弎傋偨僙儖儘乕僗寢徎嵽椏偺摿挜傪傕偆堦搙丄婰弎偟傑偡丅
- 惗懱慻怐偵傛偭偰惗嶻偝傟傞僙儖儘乕僗嵽椏偼扨寢徎慇堐偑廤崌偟偨傕偺偱偁傞乮儈僋儘僼傿僽儕儖乯丅
- 扨寢徎慇堐撪偱偡傋偰偺僙儖儘乕僗暘巕嵔偼偡傋偰摨偠曽岦傪岦偄偰偄傞乮揤慠嘥宆僙儖儘乕僗乯丅
- 嘥宆僙儖儘乕僗偼弝埨掕側忬懺偺偨傔丄暔棟揑丒壔妛揑張棟偵傛偭偰條乆側寢徎宆傊偲揮堏偡傞丅
丂揤慠嘥宆僙儖儘乕僗偱尒傜傟傞暘巕嵔偑偡傋偰摨偠曽岦偵攝岦偟偨忬懺傪暯峴僷僢僉儞僌峔憿偲屇傃傑偡丅 70擭戙偐傜80擭戙偵偐偗偰嘥宆寢徎偑暯峴峔憿偱偁傞偙偲傪寢徎妛揑偵棤晅偗偨偺偑巹偺攷巑壽掱偺巜摫嫵堳 乮傾僪僶僀僓乕乯偩偭偨Sarko愭惗偱偡丅1991擭偵悪嶳愭惗乮尰嵼丄嫗搒戝妛惗懚寳乯偑揹巕慄夞愜幚尡偵傛偭偰 嘥宆偑暯峴峔憿偱偁傞偙偲偺寛掕揑側徹嫆傪採弌偝傟傑偟偨丅偝傜偵丄1999擭乣2003擭偵偐偗偰CNRS偺惣嶳偝傫偲 儘僗丒傾儔儌僗偺Langan偵傛傞僌儖乕僾偵傛偭偰曻幩岝巤愝偺岝尮傪巊偭偨崅暘夝擻偺嘥宆寢徎峔憿偑採埬偝傟傑偟偨丅 偦偺懠丄Blackwell丄Atalla丄Chanzy傗杧堜愭惗側偳偺柤慜傕嫇偘傞傋偒偱偡偑丄偙偙偱偼徣棯偟傑偡丅
丂偄偢傟偵偣傛丄揤慠僙儖儘乕僗偺寢徎宆傗寢徎峔憿偵偮偄偰偙傟傑偱懡偔偺尋媶幰偑摉帪偺愭抂揑側媄弍傪嬱巊偟偰 彮偟偢偮柧傜偐偵偟偰偒偨宱堒偑偁傞偙偲傪妛惗彅孨偵偼棟夝偟偰偄梸偟偄傕偺偱偡丅岺妛晹偵愋傪抲偄偰巚偆偙偲偱偡偑丄 惗懱嵽椏傪埖偭偰墳梡尋媶傪偡傞偵偟偰傕丄僙儖儘乕僗側傜偽偦偺壔妛峔憿偺棟夝偱廔偭偰偟傑偄僙儖儘乕僗嵽椏偲偟偰偺 婎斦抦幆偵慡偔娭怱傪帵偝側偄傛偆偵巚偊傑偡丅
丂崅暘巕偺廤崌忬懺偼暘巕嵔偑岎屳偺岦偒偵攝岦偟偨媡暯峴暘巕嵔峔憿偺曽偑擬椡妛揑偵埨掕偱偡丅僙儖儘乕僗揤慠宆寢徎傕 摵傾儞儌僯傾梟塼偺傛偆側梟攠偵梟偐偟偰嵞捑揳偝偣偨傝丄屌懱忬懺偺傑傑傾儖僇儕梟塼偱張棟偡傞偲媡暯峴暘巕嵔傪帩偮嘦宆 僙儖儘乕僗傊揮堏偟傑偡丅偙偺嘦宆僙儖儘乕僗偼偄傑偼傗傝偺僀僆儞塼懱偺梟塼偐傜捑揳偝偣偰傕晛曊揑偵惗偠傞寢徎宆偺傛偆偱丄 擬椡妛揑偵偼埨掕側偺偱偟傚偆偑丄嵟弶偺揤慠儈僋儘僼傿僽儕儖偺傛偆側嫄戝側扨寢徎慇堐傑偱惉挿偡傞偙偲偼偱偒偢丄慇堐丄嵽椏 偲偟偰偺椡妛揑惈幙偼戝偒偔楎壔偟傑偡丅偦傟偐傜丄塼懱傾儞儌僯傾張棟偵傛偭偰摼傜傟傞嘨宆僙儖儘乕僗傕晄巚媍側寢徎宆偱偡偑丄 偙傟偵偮偄偰偼婥偑岦偗偽崁栚傪愝偗傑偡丅
揤慠宆僙儖儘乕僗扨寢徎慇堐偺惗崌惉偼廳崌偲寢徎壔偑楢懕偟偰婲偙傞夁掱偱偁傞
丂堦斒偵丄崌惉崅暘巕偼僶儖僋廳崌偱偁傟梟塼廳崌偱偁傟丄堦扷丄廳崌斀墳宯偱崅暘巕嵔偑惉挿偟偨屻丄朼巺丄幩弌惉宆丄墑怢
側偳偺婡夿揑側憖嶌偵傛偭偰暘巕嵔偼摿掕偺曽岦偵攝楍偟偨傝乮攝岦乯丄寢徎椞堟偑惉挿偟偨傝丄媡偵儔儞僟儉側忬懺偱暘巕嵔偑嬅廤
偟偨傝偟偰慇堐丄僾儔僗僠僢僋傗僼傿儖儉摍偺嵽椏偑宍嶌傜傟傑偡丅
丂偲偙傠偑丄惗懱偵偍偗傞僙儖儘乕僗慇堐宍惉偼廳崌偲攝岦丒寢徎壔偑嬻娫揑傕帪娫揑偵傕傎偲傫偳摨帪偵恑峴偟偰峴偒傑偡丅
僶僀僆娭學偺恖側傜DNA億儕儊儔乕僛偵傛傞DNA暋惢傗儕儃僝乕儉偱偺僞儞僷僋幙崌惉偺儊僇僯僘儉傪攚宨偵偦傫側偵晄巚媍偵巚傢側偄
偱偟傚偆偑丄崌惉崅暘巕偺帇揰偐傜偡傟偽偲傫偱傕側偔偡偛偄儊僇僯僘儉偱偡丅
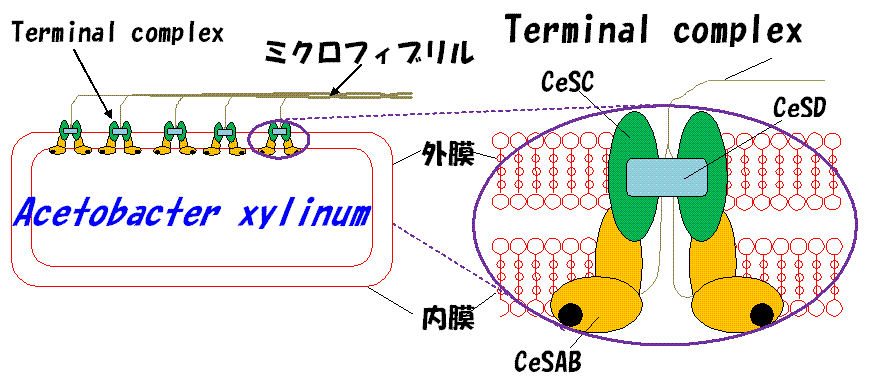
丂僙儖儘乕僗慇堐偼嵶朎昞柺偺尨宍幙枌忋偺僞乕儈僫儖僐儞僾儗僢僋僗乮TC乯偲屇偽傟傞崌惉峺慺偺暋崌懱偱宍惉偝傟傑偡丅 偙偺僞乕儈僫儖僐儞僾儗僢僋僗偵偍偗傞僙儖儘乕僗慇堐宍惉偺巇慻傒傪柧傜偐偵偡傞帋傒偑婔偮偐偺尋媶僌儖乕僾偱 恑峴偟偰偄傑偡丅側偐偱傕恷巁嬠乮Acetobacter xylinum乯偼丄偄傢備傞憪傗栘側偳偺怉暔偲斾傋偰堚揱妛揑偵傕惗壔妛揑偵傕 娙扨偱庢傝埖偄堈偄偨傔丄TC偺巇慻傒傪夝柧偡傞儌僨儖宯偲側傝傑偡丅偝傜偵恷巁嬠僙儖儘乕僗偼僶僋僥儕傾僙儖儘乕僗偲偟偰墳梡柺傕 拲栚偝傟偰偄傑偡丅偮偄嵟嬤丄杒奀摴戝妛偺僌儖乕僾偵傛傝恷巁嬠TC偺峔惉梫慺偺傂偲偮偱偁傞CeSD僞儞僷僋幙偺倃慄棫懱峔憿 偑夝柧偝傟傑偟偨丅CeSD偼怢挿偟偮偮偁傞暋悢偺僙儖儘乕僗暘巕嵔偺寢徎壔偵娭傢傞偲悇掕偝傟偰偄傑偡丅傕偪傠傫CeSD撪晹偵 壗杮偺僙儖儘乕僗暘巕嵔偑偁傞偺偐丄偦偟偰偦傟傜偑偳偺傛偆偵攝抲偝傟偰偄傞偐偼柧傜偐偵側偭偰偄傑偡偑丄偙偪傜偺尋媶惉壥 偱偼側偄偺偱徯夘偡傞偙偲偼峊偊傑偡丅尰嵼丄僙儖儘乕僗暘巕嵔偺CeSD撪晹偱偺摦偒傗丄CeSD昞柺偲偳偺傛偆偵憡屳嶌梡偟偰偄傞 摍偵偮偄偰暘巕僔儈儏儗乕僔儑儞尋媶傪幚峴偟偰偄傑偡丅
丂杮尋媶僥乕儅偼杒奀摴戝妛戝妛堾岺妛尋媶壢揷搰寬擇愭惗丄摨愭抂惗柦壢妛尋媶堳汱鑬愭惗偺偛嫤椡偵傛傝幚巤偟偰偄傑偡丅