アップル社の前CEOであったSteve Jobsによる、2005年のStanford大学卒業式における講演は歴史的なものとして大変有名です。筆者も何度か、講義の英語教材として利用しました。 講演の締めくくりに"Stay hungry, stay foolish"という言葉が引用されています。さらに"connecting the dots"という表現があり、これは、過去の様々局面で体験したり学習したことが、将来、予期しないことに繋がるという意味かと思います。それは偶然か必然、どちらを意味するかはわかりませんが。
学位研究を含めて初期の専門分野は多糖類結晶の結晶構造解析でしたが、その対象の多くがマンナン、キチン、キトサンのようなβ1,4結合を持ち、2回らせん分子鎖構造を形成する多糖類でした。2重らせんや3重らせんを構成するようなアミロースやカードランと比べて、”どうも地味だな”というのが正直なところでした。これらの2回らせん多糖分子鎖が示す結晶構造には、①分子鎖同士が水素結合によって連結され平面状または波状の分子鎖シートを形成する、②隣りあう分子鎖同士が繊維軸方向に関して同じ位置にあるか、繊維軸長のほぼ1/4だけ相対的にずれている、という特徴が挙げられます。もちろん、それらの特徴を持つ典型的な事例がセルロース結晶多形の結晶構造です。特に、繊維軸方向に関する分子鎖位置の相対的ずれ(chain staggering)については、博士論文の最終章にnon staggeringか1/4 staggringに分類される旨の総括めいた議論をしました。
いよいよ本題ですが、2011年頃、セルロースⅢI型結晶モデルの分子シミュレーション研究からの展開として、また、当時の大学院生の宇都君からの希望もあり、密度汎関数(DFT)計算への展開を考えていました。古典力学計算と異なり、DFT計算では扱える原子数が限定されるため、先行研究では4分子程度のセルロースオリゴマーを結晶格子の4隅に配置したクラスター計算結果が報告されていました。しかし、これをⅢI型結晶に適用して、水素結合がどうなった、ああなったと議論しても何ひとつ面白くありません(いわゆる銅鉄研究)。そこで思いついたのが、セルロース結晶構造を特徴づける分子鎖シートを対象としてDFT最適化計算を行ったらどうなるだろうか?というアイデアです。
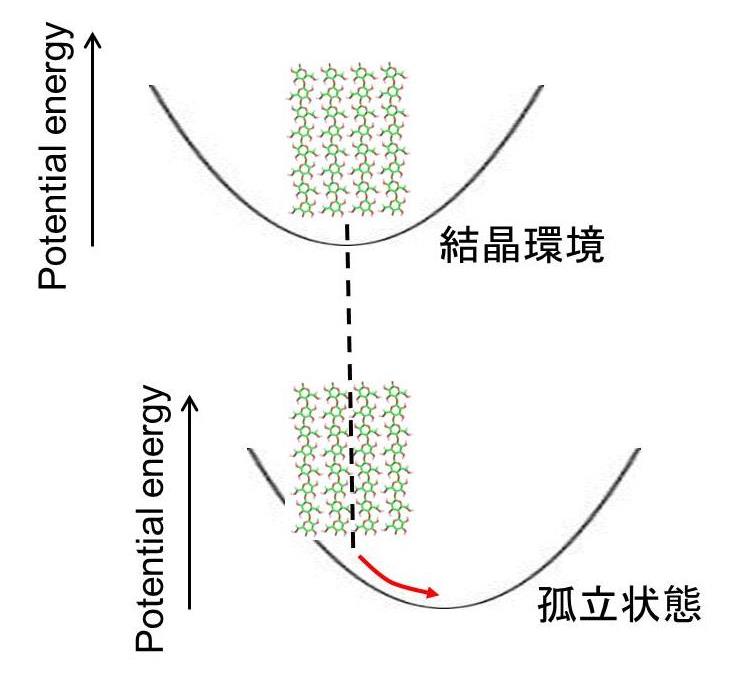 右図のように、分子鎖シートは結晶構造環境のもとでシート状態が維持されポテンシャル曲面の最安定状態にあるはずです。当然、孤立状態に置いてやると別のポテンシャル極小点へと立体構造を変化させるものと予想されます。ものの性質を調べるときは、それに刺激を与えたり、特定の要因を排除して、その影響を評価します。分子鎖シートだけ取り出すというような操作は現実には不可能なため、計算化学だからこそ可能なアプローチと言えます。当然、分子鎖シート構造は最適化の過程で初期構造から大きく構造を変化させると予想されますが、古典力学計算と異なり、電子構造の緩和を伴うDFT計算を適用することで信頼性の高い最適化計算結果が期待できます。従来、結晶を対象とする計算化学研究では、実際の結晶構造を再現することを目標とするため、この考え方は論文投稿時にレフェリーだけでなくエディターにも理解されなかった節があります。なによりも、まともな精神の持ち主であれば、最適化効率の低いDFT計算をこのような乱暴な目的に使用しません(アホみたいな回数の最適化過程の繰り返しを必要とします)。さらに、要領と常識に囚われた学生であれば、”シート構造が壊れるだけで、何の意味もないですよ!”と拒否したかもしれませんが、念願のDFT計算研究ができるということで担当の宇都君は喜々として計算に取り組んでくれました。また、彼が限られた予算内で高性能のPCクラスターを構築してくえrたため、このような負荷の大きな計算が可能になっとことも述べておきます。
右図のように、分子鎖シートは結晶構造環境のもとでシート状態が維持されポテンシャル曲面の最安定状態にあるはずです。当然、孤立状態に置いてやると別のポテンシャル極小点へと立体構造を変化させるものと予想されます。ものの性質を調べるときは、それに刺激を与えたり、特定の要因を排除して、その影響を評価します。分子鎖シートだけ取り出すというような操作は現実には不可能なため、計算化学だからこそ可能なアプローチと言えます。当然、分子鎖シート構造は最適化の過程で初期構造から大きく構造を変化させると予想されますが、古典力学計算と異なり、電子構造の緩和を伴うDFT計算を適用することで信頼性の高い最適化計算結果が期待できます。従来、結晶を対象とする計算化学研究では、実際の結晶構造を再現することを目標とするため、この考え方は論文投稿時にレフェリーだけでなくエディターにも理解されなかった節があります。なによりも、まともな精神の持ち主であれば、最適化効率の低いDFT計算をこのような乱暴な目的に使用しません(アホみたいな回数の最適化過程の繰り返しを必要とします)。さらに、要領と常識に囚われた学生であれば、”シート構造が壊れるだけで、何の意味もないですよ!”と拒否したかもしれませんが、念願のDFT計算研究ができるということで担当の宇都君は喜々として計算に取り組んでくれました。また、彼が限られた予算内で高性能のPCクラスターを構築してくえrたため、このような負荷の大きな計算が可能になっとことも述べておきます。
最初にⅢI型由来の分子鎖シートを選択したことが幸運でした。2種類の分子鎖シートのうち、(1-10)分子鎖シートは、単独でも初期構造をほぼ維持する一方で、(100)分子鎖シートへ分子鎖間でペキペキと折りたたまれるという最適化挙動が観察されました。対象計算として実施したIβ型結晶由来の分子鎖シートでは、DFT最適化によって右巻きにねじれ変形を与えました。このように、三者三様の結果が得られるとは全くの想定外です。天然セルロース結晶の右巻きねじれ変形挙動については、欧米の研究グループによってその立体化学的要因が結晶構造に基づいて議論されていましたが、この結果を得たときは、”なんだ、シート単独でもねじれるじゃないか”という感想でした。(関連論文)
ここで話は変わりますが、2012年に本学科を退職された木島剛名誉教授はScience誌を含めた多くの一流学術雑誌に業績を残された先生です。なかでもAngewandte誌に掲載された白金ナノチューブ合成は、国内外の注目を受けた成果です。何故か、木島先生はご自身が指導された博士課程学生の副審査委員に筆者を2度ほどご指名され、”あんまり関係ないな~”と思いで金属ナノ構造体の論文を読んでいたのを記憶しています。
再びⅢI型由来分子シートに戻りますが、当初、分子鎖数3本の(100)分子鎖シートが折りたたまれることが分かったので、4本シートでの再確認を宇都君にお願いしていました。彼の記憶によると2012年の4月頃だったようですが、研究室に行くと、画面の前で”先生、こんなものが出ましたけど”というような雰囲気で構造最適化のアニメーションを見せてくれました(彼が、予め、かなり苦労して作成したアニメーションでした)。それは、分子鎖シートがロール状に折りたたまれ、まさにチューブ構造が形成される様子であり、数年前まで何度もインプットされたナノチューブというキーワードが目前に示されていました。得られたチューブ構造をよく見ると隣り合う分子鎖が繊維軸に沿ってほぼ1/4ずれていることが明らかになり、博士課程時代に、散々マンナン等の結晶構造解析でお世話になった1/4 staggring構造です。従って、この簡単な規則にもとづいて分子鎖の長さや本数を拡張できることになります(ちなみに、初期構造の(100)分子鎖シートはnon-staggering)。また、ⅢI型結晶構造の特徴から平行分子鎖配置を持つチューブ構造が得られましたが、逆平行分子鎖配置も考えられます。存在しないセルロースの高次構造が対象のため、こりゃ何でも言ったもん勝ちということで、CelNTという名称まで考案してしまいました。(関連論文)
CelNTで一旦、中断しましたが、本来、ⅢI型結晶モデル研究の補足計算として実施した分子シートのDFT計算のアイデアをさらに体系的に展開しようということで、天然ⅠαとⅠβ型、ⅢI型、およびⅡ型結晶構造由来の6種類の分子鎖シートについて、より大きな系とより高精度の条件のもとで、最適化挙動を評価しました。幸い天然Ⅰ型とIIII型分子鎖シートについては、以前の結果を再現しました。Ⅱ型結晶由来の分子鎖シートでは、まず、逆平行分子鎖配向の(110)分子鎖シートはあっさりシート構造を完全に失いました。続けて、隣り合った平行分子鎖配向を持つ(010)と(020)分子鎖シートは互いに逆のキラリティにねじれ変形しました。(関連論文)
以上の孤立分子鎖シートの立体構造最適化挙動がどのような意味を持つのかははっきりとは断定できません。論文にはいろいろ議論をしましたが。レフェリーからは、結晶化のプロセスを示唆しているのは?との指摘もありましたが、そこまで解釈を広げるわけにはゆかないと思います。